|
|||||||||||||
|
|||||||||||||
|
Защита от излучения всевозможной электронной техники, Wi-fi, вышек мобильной связи, геопатогенных зон
Одним из техногенных факторов, влияющих на здоровье человека, является широкое использование различной электронной техники. Действительно, представить современную жизнь без телевизора, компьютера или мобильного телефона достаточно сложно. Но как влияют на нас эти блага цивилизации? Считается, что вред здоровью человека наносят электромагнитные поля, излучаемые мониторами персональных компьютеров, телевизорами, мобильными телефонами, радиотелефонами и прочей электронной техникой. Исследования последних лет показали, что, кроме электромагнитного излучения, есть еще и торсионные поля, которые часто называют информационными (наиболее известными исследователями в этой области являются Г.И.Шипов, А.Е.Акимов, А.В.Бобров, В.П.Казначеев, А.Р. Павленко). Экспериментально установлено, что электромагнитные излучения имеют торсионную (информационную) компоненту. Несмотря на все достижения современной фармакологической медицины, целый ряд болезней победить или существенно снизить их количество не удаеться, в связи с чем возрастает значение профилактики заболеваний, в том числе вызванных влиянием техногенных факторов. В восточной медицине здоровье человека рассматривается как гармония трех тел: физического, энергетического (биополя) и информационного. Болезни физического тела рассматриваются как результат изменений в информационном и энергетическом телах. Торсионная (информационная) компонента электромагнитных излучений действует на информационное и энергетическое тела человека, поэтому мы говорим об энергоинформационном воздействии, энергоинформационной защите. Наша компания занимается исследованием влияния торсионных полей на здоровье человека и разработкой методов защиты человека от их воздействия уже более 20 лет. Согласно исследованиям (в которых принимали участие не только специалисты нашей компании, но и специалисты из Франции, России, Швейцарии) в ряде случаев именно торсионные (а не электромагнитные) поля являются основным фактором негативного влияния на здоровье человека. В связи с чем были разработаны устройства для защиты от излучений - Форпост-1, Спинор, Vernada, Vernada Geo, Сплиттер и Вернада-авто Воздействие на человека монитора ПК
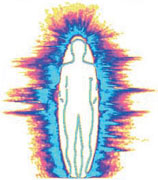
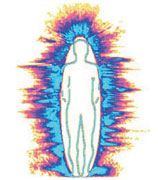
На левом снимке изображена аура человека, не подвергавшегося воздействию излучений монитора ПК (телевизора и проч.) и мобильного телефона в течении нескольких часов. Аура испытуемого имеет достаточно большую площадь, отсутствуют разрывы ауры. На правом снимке изображена аура человека, находившегося перед монитором ПК в течение 15 минут. Площадь ауры испытуемого значительно уменьшилась (энергетическое поле уменьшилось), появились разрывы ауры (полоска синего свечения либо отсутствует, либо очень тонкая).
Комплексная программа энергоинформационной безопасности |
||||||||||||
|
|||||||||||||
|
спинор, форпост, торсионные поля, аура, forpost, spinor, здоровье человека, защита, излучение, защита от излучений, биополе, безопасность, купить Спинор, Форпост-1, Vernada Geo - Spinor.in.ua, информационное, электромагнитное, компонента, нейтрализация, волны, GSM телефон, связь, базовые станции, мобильный телефон, биозащита, биологическое воздействие, |
|||||||||||||

 Все большее число людей в современном мире начинает заботиться о своем здоровье. А поскольку мы живем в век высоких технологий и технического прогресса, растет интерес к вопросу о влиянии техногенных факторов на здоровье человека.
Все большее число людей в современном мире начинает заботиться о своем здоровье. А поскольку мы живем в век высоких технологий и технического прогресса, растет интерес к вопросу о влиянии техногенных факторов на здоровье человека.